NEJM
Prespective
|
Judith Korner, M.D., Ph.D., and Rudolph L. Leibel, M.D.
New insights into the complex metabolic circuitry of energy homeostasis
have refined our understanding of the pathophysiology of obesity.
Some of these insights regarding energy homeostasis are based on the
identification of new functions for peptides that were discovered
decades ago (see Figure). It is now known that ![]() -melanocyte�stimulating
hormone, a peptide derived from proopiomelanocortin and long
recognized for its role in skin pigmentation through activity at the
melanocortin 1 receptor (MC1R), decreases food intake and increases
energy expenditure through interaction in the hypothalamus with
another melanocortin receptor, MC4R. In mice, targeted deletion of
the Mc4r gene results in obesity, hyperphagia, and
hyperinsulinemia and reduces energy expenditure. Deletion of Mc3r,
the gene encoding a melanocortin receptor that is highly expressed in
the hypothalamus and limbic system, also results in increased
adiposity due to decreased energy expenditure but does not cause
hyperphagia.
-melanocyte�stimulating
hormone, a peptide derived from proopiomelanocortin and long
recognized for its role in skin pigmentation through activity at the
melanocortin 1 receptor (MC1R), decreases food intake and increases
energy expenditure through interaction in the hypothalamus with
another melanocortin receptor, MC4R. In mice, targeted deletion of
the Mc4r gene results in obesity, hyperphagia, and
hyperinsulinemia and reduces energy expenditure. Deletion of Mc3r,
the gene encoding a melanocortin receptor that is highly expressed in
the hypothalamus and limbic system, also results in increased
adiposity due to decreased energy expenditure but does not cause
hyperphagia.
|
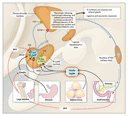
Although susceptibility to common obesity is apparently polygenic, mutations in MC4R are found in approximately 1 to 7 percent of humans whose body-mass index (the weight in kilograms divided by the square of the height in meters) is more than 40 and who become severely obese before the age of 10 years. Antagonism of anorexigenic (appetite-suppressing) melanocortin signals is caused by orexigenic (appetite-stimulating) peptides such as agouti-related protein and neuropeptide Y, which are coexpressed in a different subgroup of neurons within the hypothalamus. Agouti-related protein antagonizes the interaction between
Circulating concentrations of leptin and insulin influence these central mechanisms that control food intake and energy expenditure. The concentration of leptin in the blood is highly correlated with total fat mass; obese persons have high concentrations of leptin. Excess body fat that results in increased leptin production may actually be a correction for primary or secondary impairment of leptin-induced signal transduction in the hypothalamus. The decrease in body fat that occurs with diet-induced weight loss causes leptin concentrations to decrease and triggers responses that aim to conserve body fat. Ultimately, body-fat mass reflects the long-term balance between energy expenditure and energy intake. The latter appears to have the predominant role in maintaining this balance.
How, then, do we decide when and how much to eat? Long-term signals associated with body-fat stores are provided by leptin and insulin. These circulating molecules also modulate short-term signals that determine meal initiation and termination. Signals that provide short-term information about hunger and satiety include gut hormones, such as cholecystokinin, ghrelin, and peptide YY3�36 (PYY), and signals from vagal afferent neurons within the gastrointestinal tract that respond to mechanical deformation, macronutrients, pH, tonicity, and hormones. Neural and humoral signals are then integrated in specific regions of the hypothalamus and brain stem.
In this issue of the Journal (pages 941�948), Batterham et al. report a study of PYY, a peptide that is secreted postprandially, in proportion to the calories ingested, by endocrine L cells lining the distal small bowel and colon. The investigators found that a single infusion of PYY, as compared with an infusion of saline, reduced appetite and food consumption by approximately 30 percent at an all-you-want-to-eat buffet lunch provided two hours after the infusion. In the obese subjects, the endogenous postprandial PYY response was diminished as compared with that in the lean subjects, even though the obese subjects consumed a greater number of calories. The PYY infusion reduced hunger in both the obese and lean groups and had no effect on subjects' reports of the palatability of the meals or their feelings of nausea.
The initial release of PYY occurs shortly after food intake, presumably through neural mechanisms, before ingested nutrients arrive in the distal portion of the small intestine and the colon. The subsequent release of PYY is stimulated by nutrients, particularly carbohydrates and lipids, within the lumen of the distal portion of the small intestine and the colon. PYY decreases food intake through inhibition of gut motility (acting as an "ileal brake" to cause a sense of satiety) and by way of vagal afferent neurons that ascend from the gastrointestinal tract to the hindbrain and interactions with humoral receptors in the hypothalamus. In studies in animals, PYY inhibited the hypothalamic neuropeptide Y�expressing neurons and agouti-related protein�expressing neurons through inhibitory neuropeptide Y2 receptors, thereby disinhibiting adjacent proopiomelanocortin�expressing neurons and decreasing food intake.
The study by Batterham et al. also shows that infusion of PYY decreases fasting concentrations of the orexigenic peptide ghrelin. Ghrelin is a 28-amino-acid, acylated peptide secreted by oxyntic cells in the stomach fundus. Ghrelin acts on growth hormone secretagogue receptors to increase the release of growth hormone from the pituitary. Recently, the putative roles of ghrelin in energy homeostasis and, in particular, premeal hunger and meal initiation have been identified. Circulating ghrelin concentrations increase preprandially and decrease postprandially. Ghrelin increases food intake through the stimulation of ghrelin receptors on hypothalamic neuropeptide Y�expressing neurons and agouti-related protein�expressing neurons. Although PYY infusion reduces the concentrations of ghrelin in lean and obese subjects who are fasting and diminishes the preprandial rise in ghrelin in lean subjects, the extent to which suppression of ghrelin secretion contributes to a PYY-mediated reduction in food intake is unclear.
If ghrelin signals hunger and PYY signals satiety, can these hormones be manipulated therapeutically? Gene-knockout studies in mice reveal that one cannot easily fool the homeostatic mechanisms that maintain body fat: experimental knockouts of the ghrelin gene, AgRP, and the neuropeptide Y gene (Npy) and a double knockout of AgRP and Npy are not associated with any obvious effects on energy metabolism or food intake. In contrast, inactivating mutations of POMC, the genes that encode leptin and the leptin receptor, and MC4R produce profoundly obese phenotypes in mice as well as in humans. It appears that orexigenic pathways are so critical to survival that the absence of one peptide is compensated for by the actions of others. Studies of gut hormones after weight loss induced by dieting or surgery have provided some clues to potential pharmacologic therapies. Weight loss by caloric restriction is associated with an increase in hunger and circulating concentrations of ghrelin. After gastric bypass surgery, hunger diminishes, circulating concentrations of ghrelin decrease, and circulating concentrations of PYY increase. Hormonal changes after bypass surgery may therefore play a part in the suppression of hunger and the long-term maintenance of reduced body weight.
Although single intraperitoneal injections of PYY decrease food intake for up to seven days in rats, the results of a single infusion in humans cannot be extrapolated to predict long-term outcomes. The use of PYY may prevent counterregulatory mechanisms from overriding the stimulation of anorexigenic pathways. However, the development of antibodies or tachyphylaxis through receptor down-regulation may limit the efficacy of prolonged PYY administration. It is unlikely that any one molecule or derivative will provide a magic bullet to induce and maintain weight loss. Successful pharmacologic treatment for obesity may be possible only by simultaneously targeting the interlocking, redundant systems that drive food intake and act to resist the loss of body fat.
Source Information
From the College of Physicians and Surgeons, Columbia University, New York.